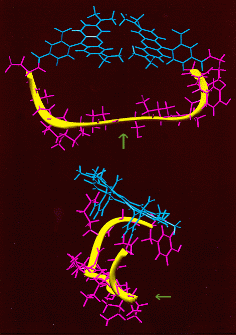
Basic Principles of Profluorescent Protease Substrates
Two different views of this peptide are shown. Note: First, the peptide backbone assumes a loop conformation similar to
1. B. Z. Packard, D. D. Toptygin, A. Komoriya, and L. Brand Profluorescenct protease substrates: intramolcular dimers described by the exciton model. Proc. Natl. Acad. Sci. (USA) 93: 11640-11645 (1996). 2. B. Z. Packard, D. D. Toptygin, A. Komoriya, and L. Brand. The design of fluorogenic protease substrates guided by exciton theory. Meth. Enzym. 278: 15-28 (1997). 3. B. Z.Packard, A. Komoriya, D. D.Toptygin, and L. Brand. Structural characteristics of fluorophores which form intramolecular H-type dimers in a protease substrate. J. Phys. Chem. B 101: 5070-5074 (1997). 4. B. Z. Packard, D. D. Toptygin, A. Komoriya, and L. Brand. Characterization of fluorescence quenching in bifluorophoric protease substrates. Biophys. Chem. 67: 167-176 (1997) 5. B. Z. Packard, D. D. Toptygin, A. Komoriya, and L. Brand. Intramolecular resonance dipole-dipole interactions in a protease substrate. J. Phys. Chem. B 102: 752-758 (1998). 6. B. Z. Packard, A. Komoriya, V. Nanda, and L. Brand. Intramolecular excitonic dimers in protease substrates: modification of the backbone moiety to probe the H-dimer structure. J. Phys. Chem. B 102: 1820-1827 (1998). On Caspase-3 nomenclature: 7. E.S. Alnemri et al. Human ICE/CED-3 Protease Nomenclature. Cell 87: 171 (1996) On DEVD-based substrate and inhibitor characteristics: 8. A. Sarin, M.-L. Wu, and P. A. Henkart. Different Interleukin-1b Converting Enzyme (ICE) family protease requirements for the apoptotic death of T-lymphocytes triggered by diverse stimuli. J. Exp. Med. 184: 2445-2450 (1996) 9. D. W. Nicholson et. al. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 376: 37-43 (1995). Intracellular Caspase-3-like activities determination using PhiPhiLux: 10. H. Hirata, A. Takahashi, S. Kobayashi, S. Yonehara, H. Sawai, T. Okasaki, K. Yamamoto, and M. Sasada. Caspases are activated in a branched protease cascade and control distinct downstream processes in Fas-induced apoptosis. J. Exp. Med. 187: 587-600 (1998) 11. J. M. Zapata, R. Takahashi, G.S.Salvesen and J.C. Reed. Granzyme release and caspase activation in activated human T-lymphocytes. J. Biol. Chem. 273: 6916-6920 (1998). 12. R.M. Siegel, D.A. Martin, L.Zheng, S.Y. Ng, J. Cohen, and M.J. Lenardo. Death-effector filaments: novel cytoplasmic structures that recruite caspases and trigger apoptosis. J. Cell Biol. 141: 1243-1253 (1998). 13. L. Guedez, W.G. Stetler-Stevenson, L. Wolff, J. Wang, P. Fukushima, A. Mansoor, M. Stetler-Stevenson. In vitro suppression of programmed cell death of B cells by tissue inhibitor of metalloproteinases-1. J. Clin. Invest. 102: 2002-10 (1998). 14. A. Komoriya, B.Z. Packard, M.J. Brown, M.-L. Wu, and P.A. Henkart. A novel intracellular caspase fluorogenic substrate for the early detection of apoptosis in thymocytes. (in preparation).
|
|
|
OncoImmunin, Inc.,
207A Perry Parkway, Suite 6
Gaithersburg, MD 20877
Phone: (301) 987-7881
FAX: (301) 987-7882
email:info@phiphilux.com
For further information, inquiries or comments, please fill out our
information request form
Copyright © 1998-2019 OncoImmunin, Inc. All rights reserved.